细胞核的功能
细胞核是细胞遗传物质储存、复制、传递及核糖体大小亚基组装的场所,在维持细胞遗传稳定性及细胞的代谢、生长、分化、增殖等生命活动中起着控制中心的作用。
一、遗传信息的贮存和复制
遗传信息是通过DNA复制、生殖细胞或体细胞分裂传递给子代或子细胞的。
(一)DNA复制是在多个复制起点上双向进行的半保留复制
真核生物中,染色体为DNA分子的载体,每条染色体为一个DNA分子,每个DNA分子上有多个复制起点。含有起点的复制单位称为复制子(replicon)。复制从复制起点开始,双向进行,在起点两侧分别形成一个复制叉(replication fork)(图8-23)。
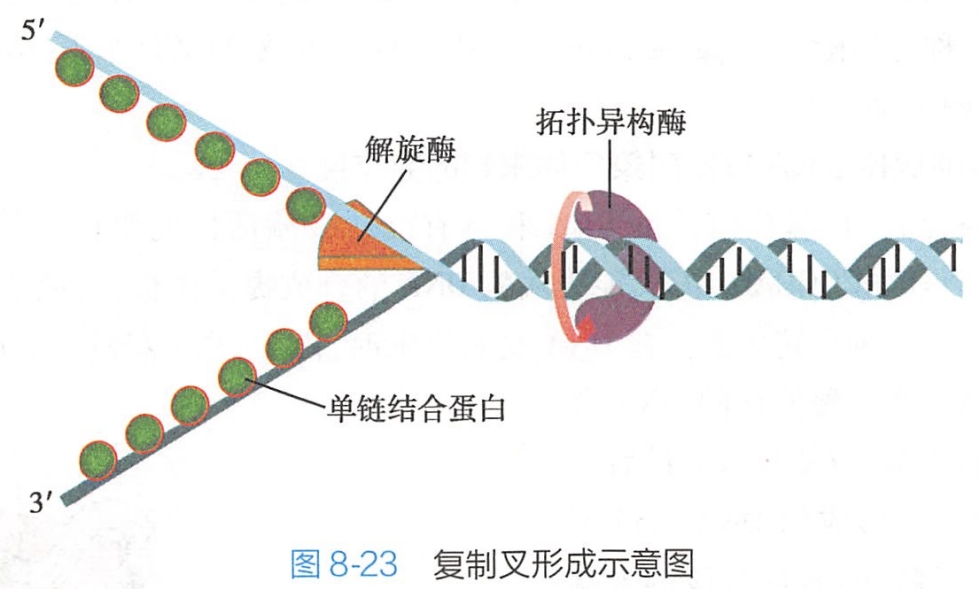
在进行DNA复制时,多个复制子可同时从起始点进行双向复制,一个复制起点的两个复制叉向两侧推进,最终将与另一起始点的复制叉相连,电镜下观察到的复制子呈一个个气泡状结构(图8-24)。
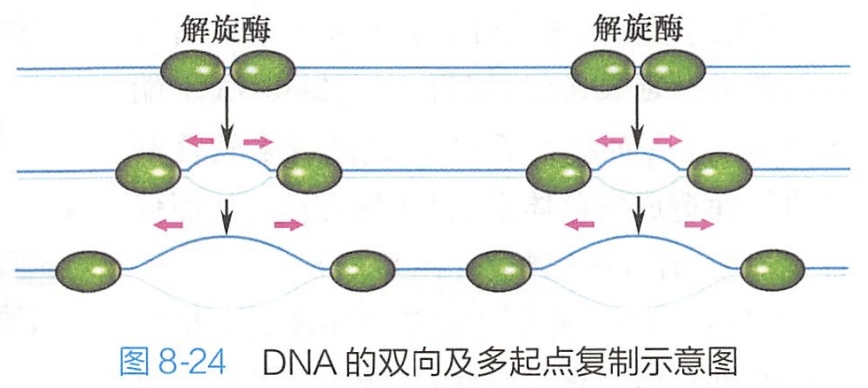
当亲代DNA分子上的所有复制子都汇合连接成两条连续的子代DNA分子时,复制得以完成。
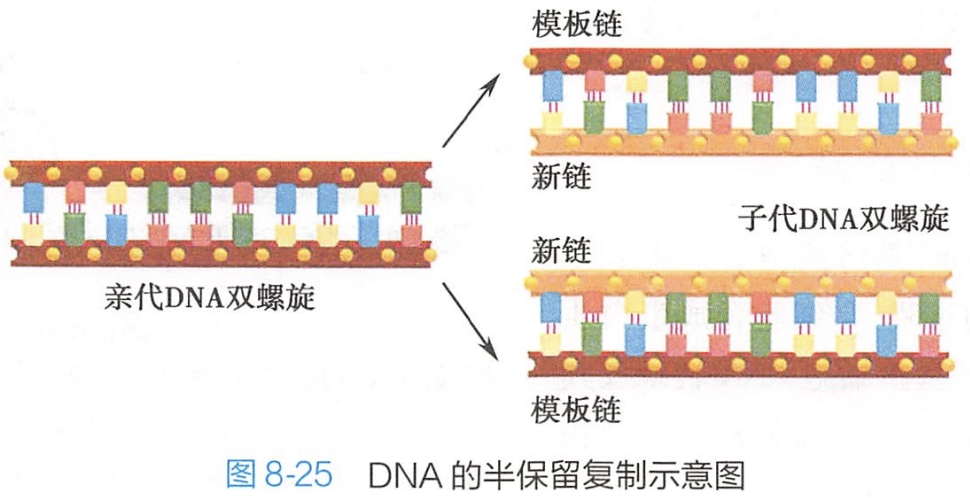
复制后的两个DNA分子中的碱基顺序与复制前的DNA分子相同,而且每一个DNA分子都含有一条旧链和一条新合成的链,因此DNA的复制是半保留复制(semi conservative replication)(图8-25)。
(二)DNA复制为半不连续性复制
由于DNA聚合酶催化合成DNA链的方向只能是5'→3',使DNA链的3'端加脱氧核苷酸,所以新合成的DNA链只能沿5'→3'的方向进行。
而DNA双链的方向一条为5'→3',另一条为3'→5',彼此反向平行。在以3'→5'方向为模板的链上,子链合成的方向与复制叉推进的方向一致,DNA是沿5'→3'方向连续复制的,速度较快,称为前导链(leading strand);
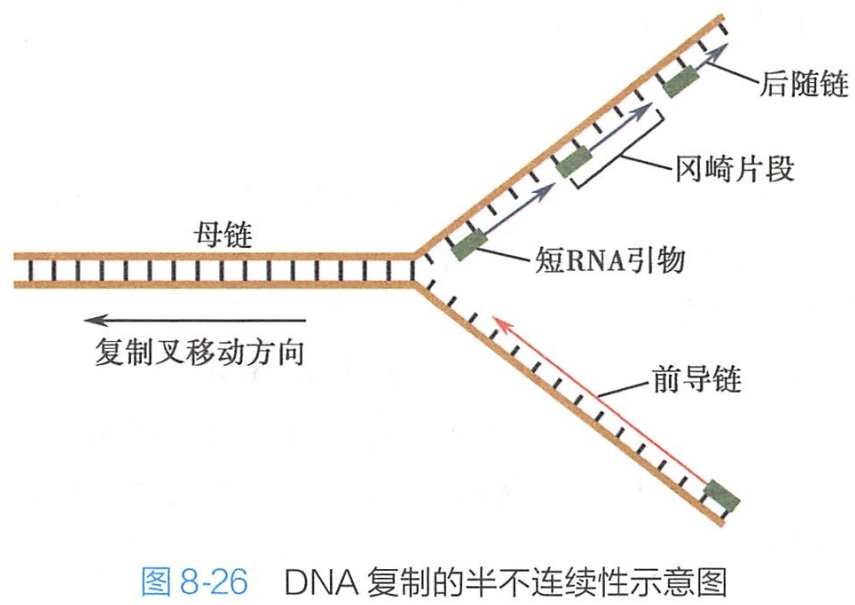
而以5'→3'链方向为模板合成的3'→5'方向的互补链,其合成方向与复制叉推进的方向相反,合成过程则需要引物(primer)的存在,即需要一个长约10bp的RNA序列以提供DNA聚合酶所需的3'端,而且每一引物只能始动合成一个100~200bp的DNA片段,称为冈崎片段(Okazaki fragment),因此在5'→3'方向的模板链上,DNA的复制是不连续的。
当一个个冈崎片段合成后,引物被去除,在DNA连接酶(DNA ligase)的作用下,补上一段DNA。所以,这一条DNA链合成较慢,称为后随链(lagging strand)。因此,DNA的复制又是半不连续复制(semi-discon- tinuous replication)(图8-26)。
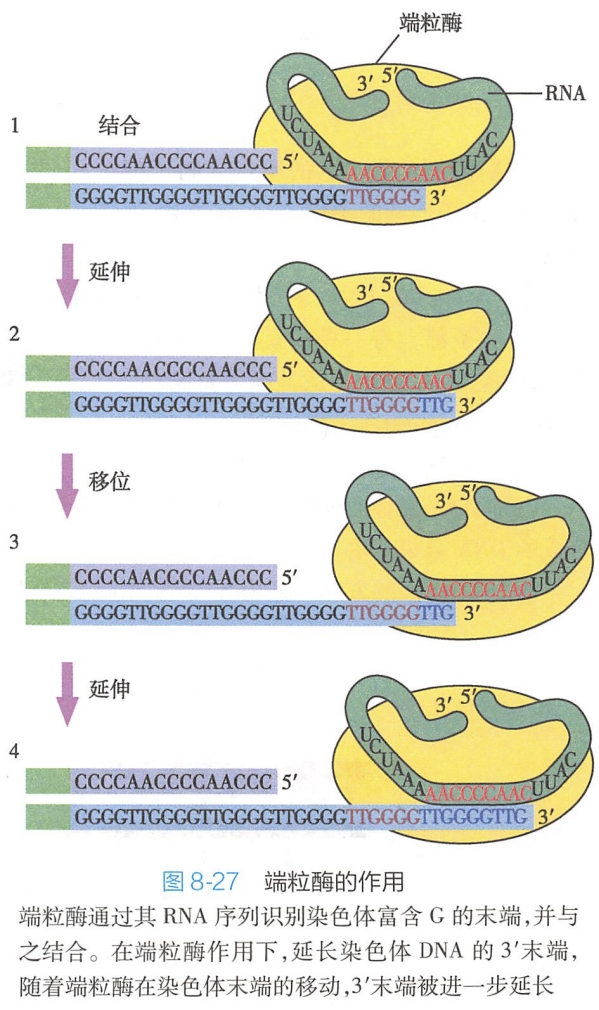
(三)端粒酶能够保持DNA复制时染色体末端的完整性
端粒是由端粒酶合成的,端粒酶是由RNA和具有反转录酶活性的蛋白质组成的复合结构,其 RNA长159bp,含一个CAACCCCAA序列,能为端粒DNA的合成提供模板,合成的方向是5'→3'。而端粒酶中的蛋白质则为一种反转录酶。
在DNA复制终末时,由于DNA双链中后随链所进行的DNA合成是不连续的,DNA聚合酶催化的DNA合成不能进行到该链的3'端(因为没那么正好,可能太短了),致使其末端最后一段序列不能进行复制,所形成的DNA新链5'端将缺失一段DNA。
端粒酶通过与该链末端的端粒序列识别并结合,以自身RNA作为模板,利用其反转录酶活性,对DNA3'末端富含G的链进行延长,通过回折,对新链DNA5'端加以补齐,从而避免了DNA链随着多轮复制的进行而造成染色体末端基因的丢失,从而保证了DNA合成的完整性(图8-27)。(实际上端粒就是用来损耗的,DNA复制得到的后随链仍然会丢失末端,不过丢失的只是端粒的一部分)
二、遗传信息的转录
转录(transcription)是将遗传信息从DNA传递给RNA分子的过程,是细胞合成蛋白质所必需的重要环节。
- RNA聚合酶I:转录的rRNA分子,在核仁部位和5SRNA以及从胞质中转运入核的核糖体蛋白结合形成核糖核蛋白颗粒(RNP),并在核仁内加工、成熟,以核糖体大、小亚基的形式转运出核;
- RNA聚合酶Ⅱ:转录的核异质RNA(heterogeneous nuclear RNA,hnRNA),首先在核内进行5'端加帽、3'端加多聚A尾以及剪接等加工过程,然后形成成熟的mRNA。由DNA转录的mRNA前体只有在核内经转录后加工修饰成为成熟的mRNA分子才能被转运出核;
- RNA聚合酶Ⅲ:5S rRNA和tRNA的转录则均由RNA聚合酶Ⅲ催化,在核内合成。
(详见第九章)